As you can probably tell from the name,
asphalt lakes are not nice places to live. Also known as tar pits, they are natural deposits of bitumen that leak up to the surface, filling the water above with all kinds of nasty substances including volcanic ash, hydrocarbons, sulphur, and metal compounds. There are only five such natural asphalt lake sites in the world, one of which is the well known
La Brea tar pits.
The largest asphalt lake in the world is
Pitch Lake on the southwest coast of Trinidad and surprisingly, it is actually home to a variety of organisms. Not just bacteria and other hardy microbes, but animals such as aquatic insects, a species of frog (
Pseudis reticulata), and some fish have also made it their home sweet home. Despite the inhospitable surroundings, there might be a perk to living in an asphalt lake. Such an harsh environment might also be intolerable for parasites, especially any external parasite which would be exposed to the asphalt-contaminated water.
 |
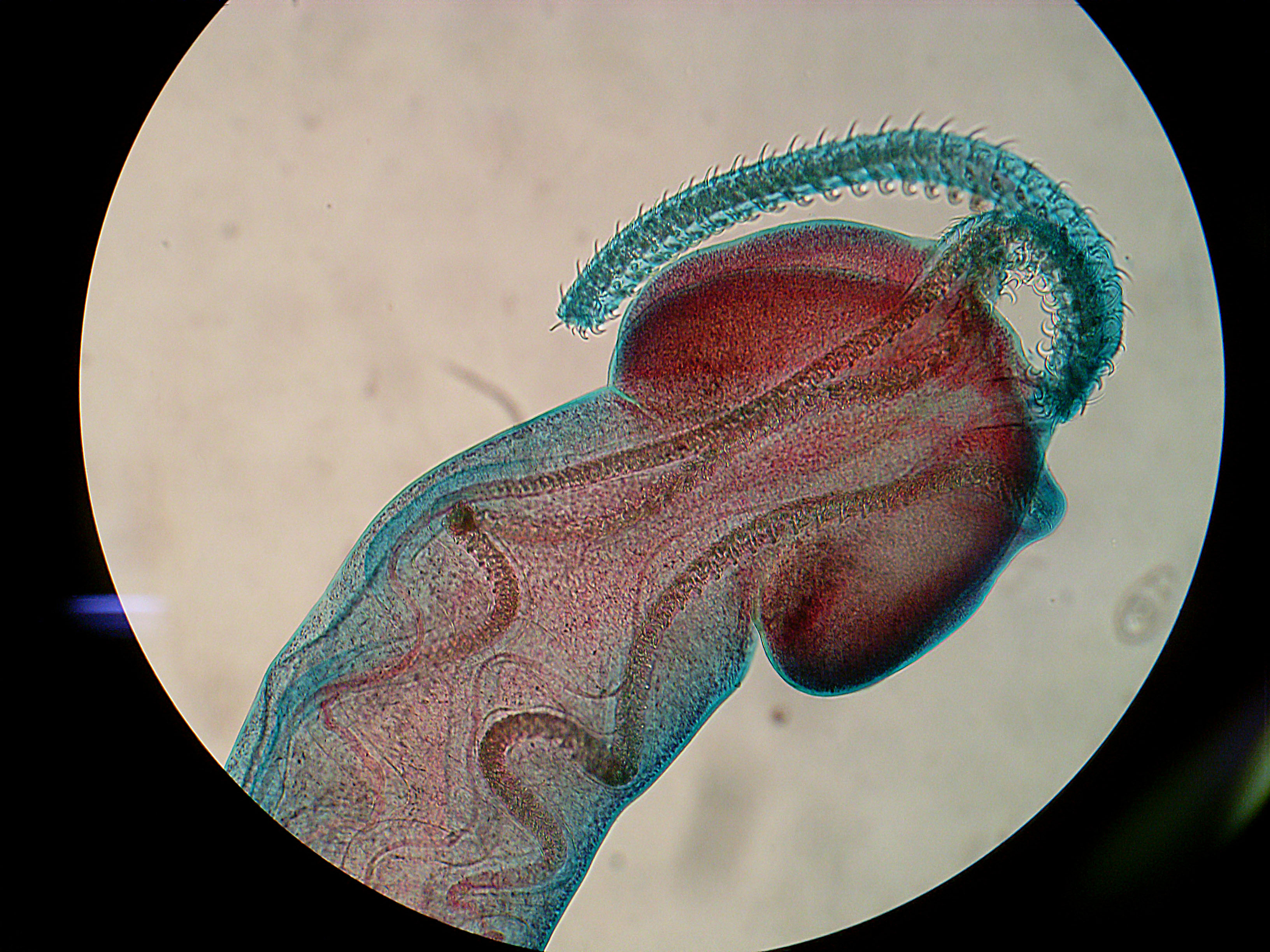
Ieredactylus rivulus
image from here |
In their natural habitat, guppies are commonly plagued by many parasites, especially ectoparasitic flatworms call monogeneans in the genus
Gyrodactylus, and in heavily infected populations as many as three-quarters of the fish will be infected. The guppies living Pitch Lake are almost completely free of parasites - except the parasites that we are featuring today -
Ieredactylus rivulus. While it is the only parasite to infect Pitch Lake guppies, it is not very abundant and they are found on fewer than five percent of the fish in any given population. Apart from Pitch Lake guppies, this parasite is only found on the giant rivulus
Anablepsoides hartii (previously known as
Rivulus hartii); another hardy inhabitant of Pitch Lake. Furthermore, the giant rivulus is also known for
wandering onto dry land every now and then, so a parasite that lives on the skin of such a fish must be pretty robust.
In the paper we are featuring today, a group of scientists conducted a series of experiments to see how the asphalt lake environment affected the guppy's parasites. In one experiment, they tested whether the Pitch Lake guppies are innately resistant to infections by placing some Pitch Lake guppies in a tank filled with dechlorinated aquarium water. Within a week, seven out of the ten guppies in the aquarium water became infested with various bacterial and fungal infection, whereas all but one of the guppies kept in the original Pitch Lake water were free from infections.
In another experiment, they tested the effect of exposure to Pitch Lake water on monogenean parasites. They collected guppies that are naturally free of monogeneans parasites from a site at the Upper Naranjo, and experimentally infected them with
Gyrodactylus by exposing them to parasite-laden guppies from the Lower Aripo, a site with high parasite prevalence. After those guppies had acquired some parasites from their infected cousins, the scientists transferred one group of the newly-infected guppies into a tank filled with water they collected from Pitch Lake that has been diluted to a quarter of its original concentration, and another group into tank of dechlorinated aquarium water. Within 48 hours, the guppies transferred into the diluted Pitch Lake water had lost their newly-acquired parasites, whereas those transferred into the aquarium water were stuck with their new parasites.
Both of those experiments showed that the Pitch Lake water was playing a key role in keeping the Pitch Lake guppies free from (most) infection, and that
I. rivulus must have some special adaptations which allows it to survive on fish swimming in a pond filled with bitumen. So if
I. rivulus can survive on asphalt lake guppies, what is to stop them from taking on guppies living in less noxious surroundings? Perhaps in the extreme environment of the Pitch Lake,
I. rivulus does not face competition from other parasites and can have the host all to itself, whereas in other guppy populations they will be competing with rapidly breeding parasite like
Gryodactylus and get shoved aside.
So while asphalt lakes might not be attractive places to live, such extreme environments can provide their inhabitants with a refuge from all but the most hardy parasites.
Reference:
Schelkle B, Mohammed RS, Coogan MP, McMullan M, Gillingham EL, van Oosterhout C, Cable J. (2012) Parasites pitched against nature: Pitch Lake water protects guppies (
Poecilia reticulata) from microbial and gyrodactylid infections.
Parasitology 139:1772-1779















{kind=link}